Жизнь во Вселенной
- Жизнь во Вселенной — под этим термином следует понимать комплекс проблем и задач, направленных на поиск жизни. В самом общем случае жизнь трактуется максимально широко — как активная форма существования материи, в некотором смысле высшая по сравнению с её физической и химической формами существования. Таким образом, в общей постановке задачи нет требования, чтобы жизнь была похожа на земную, и есть целый ряд теорий, доказывающий, что жизнь может принимать и другие формы. Однако, основной подход, использующейся в астробиологии при построении стратегий поиска, состоит из двух этапов:
Изучение возникновения жизни на Земле. Выработка основных положений. В роли скелета выступают:
Данные о геологической жизни планеты, в частности вулканизме, тектонике и магнитном поле.
Данные об истории климата и наше понимание механизмов, регулирующих его.
Основные представления об устройстве жизни, в частности о ДНК, клетках и границ выживания живых организмов
Данные о происхождении живых организмов и их эволюции.
Согласование основных положений с астрономическими наблюдениями и теориями и целенаправленный поиск. Включает в себя:
Поиск жизнепригодных экзопланет
Построение теорий формирований, включающие в рассмотрение сложные молекулярные образования, из которых впоследствии могла зародиться жизнь.
Изучение Солнечной Системы и соотнесение полученных данных с данными об экстрасолнечных системахТакже в отдельную область исследований может выделить поиск внеземных цивилизаций. Основных вопросов в данной области три:
Где искать?И здесь в построении стратегии исследований крайне важная, если не ключевая, роль принадлежит уравнению Дрейка, в дополнении с типами цивилизаций по Кардашеву.
Связанные понятия
Предметом данной статьи является современное представление об основных этапах развития Вселенной с момента её образования и до наших дней. Оно базируется на следующих теориях.
Источник
Жизнь во Вселенной
Из Википедии — свободной энциклопедии
Жизнь во Вселенной — под этим термином следует понимать комплекс проблем и задач, направленных на поиск жизни. В самом общем случае жизнь трактуется максимально широко — как активная форма существования материи, в некотором смысле высшая по сравнению с её физической и химической формами существования. Таким образом, в общей постановке задачи нет требования, чтобы жизнь была похожа на земную, и есть целый ряд теорий, доказывающий, что жизнь может принимать и другие формы. Однако, основной подход, использующейся в астробиологии при построении стратегий поиска, состоит из двух этапов [1] :
- Изучение возникновения жизни на Земле. Выработка основных положений. В роли скелета выступают [2] :
- Данные о геологической жизни планеты, в частности вулканизме, тектонике и магнитном поле.
- Данные об истории климата и наше понимание механизмов, регулирующих его.
- Основные представления об устройстве жизни, в частности о ДНК, клетках и границ выживания живых организмов
- Данные о происхождении живых организмов и их эволюции.
- Согласование основных положений с астрономическими наблюдениями и теориями и целенаправленный поиск. Включает в себя:
- Поиск жизнепригодных экзопланет
- Построение теорий формирований, включающие в рассмотрение сложные молекулярные образования, из которых впоследствии могла зародиться жизнь.
- Изучение Солнечной Системы и соотнесение полученных данных с данными об экстрасолнечных системах
Также в отдельную область исследований может выделить поиск внеземных цивилизаций. Основных вопросов в данной области три:
И здесь в построении стратегии исследований крайне важная, если не ключевая, роль принадлежит уравнению Дрейка, в дополнении с типами цивилизаций по Кардашеву. [3]
Источник
Есть ли жизнь во Вселенной?
С тех пор, как люди признали масштабность Вселенной, мы интуитивно поняли, что жизнь должна существовать где-то в нашей галактике или какой-то далекой галактике. Если Вселенная содержит миллиарды галактик, и если каждая галактика содержит миллиарды звезд, и если у части этих звезд есть планеты, подобные Земле , тогда сотни, а может быть, и тысячи инопланетных цивилизаций должны существовать во всем космосе. Правильно?
Некоторое время наука довольствовалась только логикой. Затем, в 1995 году, астрономы обнаружили первые планеты вне нашей Солнечной системы. С тех пор они обнаружили около 300 таких планет. Хотя большинство из них большие, горячие планеты, похожие на Юпитер (именно поэтому их легче найти), меньшие планеты, подобные Земле, тоже начинают проявлять себя. В июне 2008 года европейские астрономы обнаружили три планеты, все немного больше Земли, вращающихся вокруг звезд.
Эти открытия послужили подтверждением для тех, кто занимается поиском внеземной разумной жизни или SETI . Гарвардский физик и лидер SETI Пол Горовиц смело заявил в интервью журналу TIME за 1996 год: «Интеллектуальная жизнь во вселенной? Гарантированно. Разумная жизнь в нашей галактике? Вполне возможно, я бы дал вам почти любые шансы, которые вы хотели бы».
И все же его энтузиазм должен быть смягчен тем, что ученые называют парадоксами Ферми. Этот парадокс, впервые сформулированный физиком Энрико Ферми в 1950 году, задает следующие вопросы: если инопланетяне настолько распространены, почему они не побывали? Почему они не общались с нами? Или, наконец, почему они не оставили какой-то след своего существования, например, тепла или света или каких-либо других электромагнитных субпродуктов?
Возможно, внеземная жизнь не так распространена. Или, возможно, внеземная жизнь, порождающая развитые цивилизации, не так распространена. Если бы только астрономы могли количественно оценить эти шансы. Если бы они имели формулу, которая учитывала бы все правильные переменные, связанные с внеземной жизнью. Как выясняется, они это делают. В 1961 году, для организации первой серьезной конференцию по SETI, радиоастроном Фрэнком Дрейком представил формулу, теперь известную как уравнение Дрейка, которая оценивает количество потенциальных интеллектуальных цивилизаций в нашей галактике. Формула вызвала много споров, главным образом потому, что она приводит к широко варьируемым результатам. И все же это единственный лучший способ количественно определить, сколько инопланетян можно встретить. Давайте подробнее рассмотрим уравнение и его последствия.
Уравнение Дрейка
Попытка рассчитать вероятность существования внеземной жизни во Вселенной на самом деле довольно сложна. Вселенная не является статической средой. Звезды рождаются, живут и умирают. Некоторые звезды формируются вместе с планетами. Другие нет. Только некоторые из этих планет имеют правильные условия для поддержки жизни.
Жизнь — довольно сложная переменная. Некоторые планеты могут поддерживать сложные органические молекулы — белки и нуклеиновые кислоты — и больше ничего. Другие планеты могут поддерживать простые, одноклеточные организмы. И все же другие могут поддерживать многоклеточные организмы, в том числе достаточно продвинутые, чтобы развивать технологии для перемещения или отправки сигналов в космическое пространство. Наконец, даже организмы, которые очень хорошо адаптировались к окружающей среде, не навсегда выживают. Как динозавры, и Римская империя иллюстрируют здесь на Земле, что все династии заканчиваются, от катаклизмов или иначе.
Фрэнк Дрейк должен был объяснить все эти переменные при разработке формулы для количественной оценки шансов на поиск внеземной жизни. Его первой задачей было решить, что он хотел рассчитать. Во-первых, он ограничил свое мышление инопланетянами в нашей домашней галактике — и только те, которые могут быть способны к межзвездному общению. Затем он ввел математический фактор для учета всех условий, необходимых для развития таких цивилизаций. В результате получается следующая формула
В этом уравнении N — количество обнаруживаемых цивилизаций в нашей галактике. Другие переменные описаны ниже:
R — скорость образования звезд в галактике;
fp — доля звезд, образующих планеты;
ne — количество планет, гостеприимных для жизни (т. е. землеподобных планет);
fl — доля этих планет, на которых фактически возникает жизнь;
fi — доля этих планет, на которых возникает разумная жизнь;
fc — доля этих планет с разумными существами, способными к межзвездной коммуникации;
L — это время, когда такая цивилизация остается обнаруживаемой.
Единственной переменной, известной с какой-либо степенью определенности, является скорость образования звезд R. В Млечном Пути, типичной спиральной галактике, новые звезды формируются со скоростью примерно четыре в год. Астрономы чувствуют себя наиболее неуверенными в переменной L — это время, когда цивилизация остается обнаруживаемой. Для L использовались различные оценки, от 10 лет до 10 миллионов лет.
Астрономы могут дать обоснованные догадки об остальных переменных. Например, из девяти планет в нашей Солнечной системе только четыре, которые называют земными планетами, то есть те, которые имеют сплошную поверхность. Из этих земных планет только Земля поддерживает жизнь. Если взять нашу Солнечную систему как представительную, то можно утверждать, что ne равно 1/4 или 0,25. Аналогичные догадки были сделаны относительно других переменных, и, что интересно, все они имеют очень похожие значения, обычно в диапазоне от 0,1 до 1,0. Итак, типичный расчет может выглядеть так:
N = 4 × 0,5 × 0,25 × 0,2 × 0,2 × 0,2 × 3 000 000
который дает нам 12 000 цивилизаций в нашей галактике.
Первоначальные расчеты Дрейка были очень близки к этому значению для N. Когда он запустил числа, он предсказал, что в Млечном Пути может быть 10 000 обнаруживаемых цивилизаций. Карл Саган, лидер движения SETI, был еще более щедр, когда предположил, что в галактике может существовать 1 миллион цивилизаций.
Неудивительно, что астрономы были настолько оптимистичны, что в 1960-х годах они стали искать пришельцев внеземной жизни.
Тестирование и пересмотр уравнения Дрейка
Вооруженные оценкой числа коммуникативных цивилизаций в нашей галактике, ученые SETI решили их найти. У них было два основных варианта: личный контакт или сигнал из космоса. Первый сценарий требовал, чтобы инопланетяне посещали людей или наоборот. Это казалось крайне маловероятным, учитывая расстояния между нашей Солнечной системой и другими звездами Млечного Пути. Последний сценарий включал радиовещание, отправляя или получая электромагнитные сигналы в космос.
В 1974 году астрономы намеренно передали сообщение в 210 байт из обсерватории Аресибо в Пуэрто-Рико в надежде оповестить цивилизацию в глобулярном звездном скоплении М13. Сообщение содержало фундаментальную информацию о людях и нашем положении во Вселенной. Но такого рода активное общение было редкими. Астрономы в основном полагаются на пассивное общение — слушая передачи, посылаемые чужими цивилизациями.
Радиотелескоп является основным инструментом для таких экспериментов по прослушиванию, потому что он предназначен для обнаружения более длинноволновой энергии, которую оптические телескопы не видят. В радиоастрономии гигантское блюдо направляется на соседнюю солнечную звезду и настраивается на микроволновую область электромагнитного спектра. Полоса частот микроволн, между 1000 МГц и 3000 МГц, является идеальной, поскольку она менее загрязнена нежелательным шумом. Она также содержит линию излучения — 1420 МГц — что астрономы могут слышать как постоянное шипение по всей галактике. Эта узкая линия соответствует энергетическим преобразованиям, происходящим в нейтральном водороде. Как первобытный элемент вселенной, водород должен быть известен всем межгалактическим цивилизациям, что делает его идеальным маркером. Несколько команд со всего мира систематически слушают звезды Млечного Пути и соседние галактики с 1960 года.
Несмотря на их коллективные усилия, никакие поиски SETI не получили подтвержденный внеземный сигнал. Наши телескопы подобрали несколько необъяснимых и интригующих сигналов, таких как так называемый «Wow» сигнал, обнаруженный исследователями в Университете штата Огайо в 1977 году, но передача не была повторена таким образом, чтобы она дала неоспоримые доказательства внеземной жизни. Все это возвращает нас к парадоксу Ферми: если тысячи цивилизаций в галактике Млечного Пути, почему мы их не обнаружили?
Поскольку Дрейк и Саган сделали свои оценки, астрономы стали более консервативными. Пол Горовиц, который смело гарантировал существование внеземной жизни, дал более скромные результаты из уравнения Дрейка, установив, что N может быть ближе к 1000 цивилизаций.Но даже эта цифра может быть слишком большой.
В 2002 году издатель журнала «Скептик» Майкл Шермер утверждал, что астрономы не были достаточно критичны в оценке L, продолжительности существования цивилизаций. Глядя на 60 цивилизаций, которые существовали на Земле с самого начала человечества, Шермер придумал значение для L, которое варьировалось от 304,5 лет до 420,6 лет. Если вы вставляете эти числа в уравнение Дрейка, вы обнаружите, что N равно 2.44 и 3.36 соответственно. Подстройте номера еще немного, и вы можете легко заставить N упасть до одного или даже ниже. Внезапно шансы услышать сигнал от внеземной формы жизни значительно ниже.
Даже самые восторженные сторонники SETI обеспокоены отсутствием результатов, вызванных более чем 40-летним прослушиванием космических радиоволн. И все же большая часть этого поиска была ограничена нашей домашней галактикой. Даже если в галактике всего три или четыре цивилизации, есть миллиарды и миллиарды галактик. Это снова ставит вопрос о поиске внеземной жизни, поэтому многие астрономы SETI используют тот же подход к своей работе, что и лотерейные игроки: вы не можете победить, если не играете.
Источник
Что такое Жизнь во Вселенной: четыре базовых принципа вместо трёх характерных функций
Данная статья мотивирована необходимостью построить более общую картину, что такое жизнь (и какой она может быть) по отношению к остальным явлениям во Вселенной.
(Является переводом довольно любопытной научной статьи от 16 апреля 2020 года).
Введение. Зачем нужно новое определение для жизни?
Мы утверждаем, что большинство стандартных определений жизни ограничены и могут помешать будущим астробиологическим исследованиям в задаче поиска новых форм жизни. На данный момент в NASA используется такое определение — «самоподдерживающаяся химическая система, способная к эволюции Дарвина». Несмотря на то, что это довольно точное описание жизни, которая присутствует в данный момент на планете Земля, поиск во Вселенной явлений, которые соответствуют этому определению, похож на игру в дартс игроком, который концентрируется только на центре мишени. Для тех кто не знает, как устроен дартс, мы перечислим три основные проблемы при таком подходе к игре:
Попасть сложно из-за слишком маленькой целевой зоны
Это не самая выгодная зона для попадания (гораздо лучше целиться в 3х60)
Есть множество других довольно выгодных участков для прицеливания
Следовательно, при поиске внеземной жизни мы должны учитывать, что:
Жизнь, очень точно подходящая под признаки той, с которой мы знакомы, может быть очень редкой во Вселенной, однако более общий класс жизнеподобных феноменов может быть куда более распространённым.
Могут существовать системы, которые только предстоит открыть или хотя бы представить, и они будут более успешно удовлетворять критериям жизни, даже по сравнению с земной.
Ослабив наши ограничения в определении жизни, мы откроем весь спектр параметров физических и химических взаимодействий, которые могут создавать жизнь.
Так же нас мотивируют жаркие споры в области происхождения жизни, которые разделили исследователей на множество лагерей, в каждом из которых спорщики ищут свой, «единственно истинный» сценарий появления жизни. Мы утверждаем, что большая часть споров произошла из-за различий в предположении о том, что есть жизнь и, соответственно, каким было её начало.
Переопределяя сам термин «жизнь», мы надеемся, что это поможет более глубокому взаимопониманию и информационному обмену между сообществами исследователей.
Характерные функции жизни
Многие теории происхождения жизни сосредоточены на объяснении возникновения «характерных функций» — специфических аспектов современной земной биологии, которые, как предполагается, присутствовали при ее возникновении. Часто также подразумевается, что они имеют более фундаментальное значение для жизни в том смысле, что после их естественного появления должны возникать и остальные признаки жизни.
Другой взгляд предполагает существование неких «законов сохранения», подобных тем, что играют важную роль в физике и инженерии. То есть какие-то материальные и функциональные ограничения, которые присущи для жизни с самого начала. Любая система, которая не соответствует этим ограничениям не может называться жизнью (в рамках этого подхода).
Одним из примеров такого подхода является «принцип сохранения химии», на который ссылаются в [3 ]. Их аргумент выглядит таким образом: «Химические свойства организмов более консервативны, чем изменяющаяся окружающая среда, и поэтому сохраняют информацию о древних условиях окружающей среды» [4 ]. В соответствии с этим принципом поиск контекста происхождения жизни, по сути, представляет собой поиск пребиотических сред, максимально приближенных к клеточным условиям.
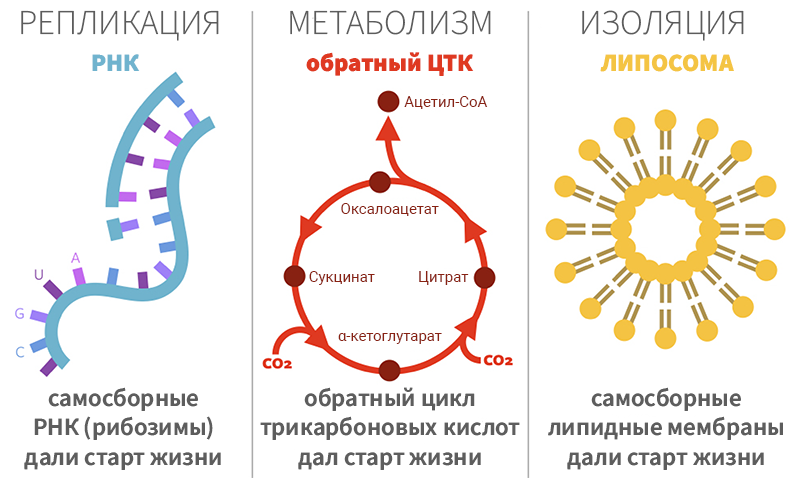 Рисунок 1. Три распространённых примера характерных функций жизни в теориях её происхождения.
Рисунок 1. Три распространённых примера характерных функций жизни в теориях её происхождения.
Примерами характерных функций являются репликация РНК по шаблонам, циклы реакций, формирующие ранние виды метаболизма и изоляция систем липидными мембранами от окружающего мира (рис. 1). Это приводит к возникновению различных «первичных» теорий возникновения жизни. Например, в так называемой «гипотезе мира РНК» считается, что «согласно теории эволюции Дарвина неживая материя должна самоорганизоваться до получения поведения, которое мы приписываем биологии»[5 ], а наступление дарвинизма произойдет с абиотическим образованием длинноцепочечных РНК-полимеров.
И наоборот, во многих «метаболических» теориях жизнь рассматривается как «лишь один из примеров самоорганизующихся систем, которые возникают по всей Вселенной как неизбежный результат термодинамических неравновесий»[6 ]. С этой точки зрения жизнь, прежде всего, служит рассеиванию (диссипации) специфических неравновесий, используя их энергию для поддержания своего низкоэнтропийного состояния [7 ,8 ].
Сосредоточение внимания на характерных функциях жизни приводит каждую из этих теорий к различным взглядам на благоприятные условия для возникновения жизни — и это является серьёзным источником напряжения в научном сообществе.
Лабораторные эксперименты показывают, что приповерхностные среды, подверженные УФ-излучению и (возможно, эпизодическому) взаимодействию с атмосферными условиями и/или снабжаемые подходящими молекулами из внешних источников, идеально подходят для синтеза РНК, например [12 ].
С другой стороны, глубоководные щелочные гидротермальные источники являются центрами фокусировки окислительно-восстановительных процессов и неравновесия рН, которые могут приводить к возникновению протометаболических циклов, например, [6 ].
Однако, геотермальные пузырьки благоприятны для спонтанной самосборки липидных везикул, что отчасти обусловлено низкими концентрациями двухвалентных катионов [13 ].
Каждая теория, пытающаяся объяснить какую-то характерную функцию, присущую земной биологии, содержит неявное предположение о том, что эта самая характерная функция присутствовала при появлении жизни и что она является основополагающей.
Однако, учитывая изначальное отсутствие ископаемых или геологических свидетельств, нет почти никаких доказательств того, что какая-либо из этих функций действительно присутствовала в начале жизни.
Основываясь на филогенетических исследованиях, наши знания о самых ранних формах жизни на Земле прекращаются на последнем универсальном общем предке (last universal common ancestor, LUCA). Такие исследования пролили свет на природу LUCA — скорее всего, это клеточное существо с хемиосмотическим метаболизмом, генетика которого была записана в ДНК/РНК. Однако, LUCA почти наверняка не являлся исходной формой жизни на Земле [14 ]. Кроме того, филогенетические реконструкции LUCA по своей природе проблематичны и потенциально ненадежны в связи с ограничениями биоинформационных техник.
Горизонт событий исследований происхождения жизни
Существует «горизонт событий», как подходящий по смыслу термин из астрофизики, в исследованиях происхождения жизни (рис. 2).
Подходы «сверху-вниз», такие как молекулярная филогенетика, используют подсказки из сохранившейся жизни, чтобы проследить историю жизни в направлении ее происхождения. Такой подход может привести нас только до LUCA.
Подходы «снизу-вверх», направленные на моделирование синтеза пребиотических молекул и/или наступление протоживых структур и функций, могут в один прекрасный день привести к созданию абиогенеза. Однако такое «пробное происхождение» не будет являться LUCA.
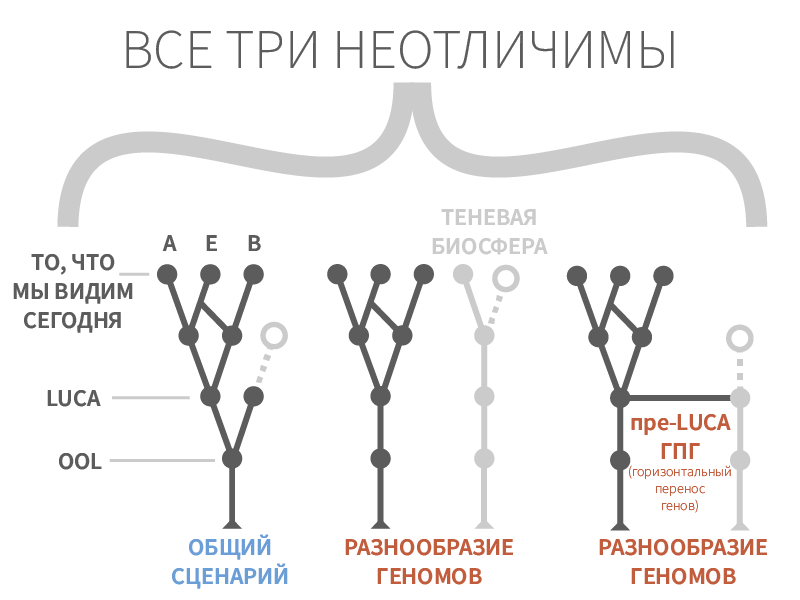 Рисунок 3. Невозможность различия сценариев происхождения жизни при подходе «сверху-вниз».
Рисунок 3. Невозможность различия сценариев происхождения жизни при подходе «сверху-вниз».
Кроме того, нисходящие подходы ограничены в своем понимании происхождения жизни, поскольку дерево жизни, которое мы видим сегодня, может быть достигнуто самыми разными теоретическими сценариями (рис. 3). Эти сценарии могут включать в себя множество никогда не сходящихся генов, в результате чего образуются совершенно разные геномные ветви, которые либо уже вымерли, либо все ещё присутствуют в качестве «теневой биосферы» [15 ].
Правдоподобные сценарии появления жизни могут включать горизонтальный перенос генов между различными геномами, которые, возможно, беспорядочно в протобиологическом рибофильме [16 ], так что LUCA на самом деле является смесью из целого множества жизненных форм — некая «последняя универсальная общая группа предков» (LUCAS) [17 ].
Если это так, то «дерево» жизни может быть даже более подходящей метафорой, чем мы могли первоначально подумать — в то же время как оно разветвляется вверх в разнообразие существ, населявших нашу планету после LUCA(S), оно также разветвляется и вниз в запутанную сеть ранних экспериментов жизни, о которых мы никогда не сможем получить сведений.
Исторический vs Синтетический vs Универсальный сценарии происхождения
Существует более глубокая проблема — исследователи происхождения жизни разных направлений, возможно, не ищут более общих объяснений. В работе [18 ] определены три различные категории описания происхождения: историческое, синтетическое и универсальное (рис. 4).
Исторические сценарии происхождения жизни целиком основаны земных наблюдениях и ограничены нашими знаниями об окружающей среде на ранних стадиях развития Земли и основаны на двух концепциях — LUCA и современное древо жизни. Все теории о «характерных функциях» являются историческими сценариями.
Универсальные сценарии описывают шаги, необходимые для абиогенеза в любых условиях. Они практически не ограничены условиями ранней Земли, траекторией развития биосферы на нашей планете или химической природой жизни в том виде, в котором мы ее знаем, — все это является лишь одним из множества возможных вариантов действия универсального сценария.
Пока нет уверенности в том, что универсальные сценарии существуют, но предпринимаются некоторые попытки объяснить функции жизни с абстрактной и фундаментальной физической точки зрения [22 ,23 ,24 ,25 ].
Следовательно, различные сценарии, разработанные исследователями, затрагивают не один, а множество научных вопросов. Одним из них обычно является происхождение жизни на Земле, которое распространяется и на генезис земноподобной жизни на других мирах.
Однако существует совершенно отдельный вопрос о происхождении «жизнеподобных» систем, как земных, так и неземных, в инопланетных средах.
Ещё один вопрос заключается в том, как мы могли бы создать полностью искусственные структуры, но которые мы бы считали живыми.
Все вопросы — будь то объяснение происхождения хемиосмотических процессов в земной жизни, избежание «катастрофы ошибок» в любой реплицирующей системе, основанной на шаблонах, или возникновение «порядка из хаоса» в целом — актуальны, но они не обязательно задают один и тот же вопрос.
Эти разрозненные цели и задачи могут привести к тому, что сообщество исследователей столкнётся с проблемой «лебедя, рака и щуки» (вместо взаимопомощи), излишних споров (вместо консенсусного обмена идеями) или апатии (вместо подлинного интереса к работам друг друга).
В этой статье вводится новый словарь и новое определение жизни в ответ на эти опасения.
Определение Y-жизни
Примечание переводчика — авторы предлагают использовать нестандартное слово для нового, более широкого, определения жизни — Lyfe (в отличии от стандартного английского «life«). Для русского языка не придумал способа как-либо приемлемо исказить привычное слово, поэтому далее будет применяться термин «Y-жизнь«.
Напишите в комментариях, может быть у вас будут более удачные варианты.
Мы стремимся переопределить сам термин «жизнь» в более широком смысле, однако не собираемся смешивать это определение с тем конкретным видом жизни, который мы видим на Земле. Мы придумали новый термин — «Y-жизнь». Отныне мы будем называть земную жизнь (такую, какой мы её знаем) «жизнью», а термин «Y-жизнь» будет являться термином с наиболее общим смыслом. Эти два обозначения различаются следующим образом:
Жизнь олицетворяет жизнь в том виде, в каком мы ее знаем; она использует специфические неравновесия и классы компонентов земной жизни. Жизнь — это автокаталитическая сеть металлоорганических химикатов в водном растворе, которая записывает и обрабатывает информацию об окружающей её среде в молекулярном виде и достигает динамического порядка за счет рассеивания любого подмножества следующих неравновесий: окислительно-восстановительные градиенты, хемиосмотические градиенты, видимые/тепловые фотоны и др.
Y-жизнь представляет собой любое гипотетическое явление во Вселенной, которое удовлетворяет фундаментальным принципам живого состояния, независимо от типов эксплуатируемых неравновесий либо компонентов. Y-жизнь представляет собой любое гипотетическое явление, которое поддерживает своё низкоэнтропийное состояние путём диссипации и конверсии термодинамических неравновесий, использует цепи автокаталитических реакций для достижения нелинейного роста и распространения, использует гомеостатические регулирующие механизмы для обеспечения стабильности и смягчения внешних возмущений, а так же собирает и обрабатывает функциональную информацию об окружающей среде.
Понятие «жизнь, какой мы ее не знаем» не ново. Тем не менее, традиционные определения жизни не позволяют строго разграничить понятия «жизнь, какой мы ее знаем» и «жизнью, какой мы ее не знаем». Их расплывчатость не позволит нам вынести какой-то определённый вердикт, если разница между двумя категориями велика.
Например, согласно определению жизни, данному НАСА (см. введение), следует ли считать самоподдерживающуюся химическую систему, которая развивается не по Дарвину, «жизнью, какой мы её не знаем» или вообще не стоит её считать жизнью?
Чтобы исправить это, мы и разработали наши критерии Y-жизни на основе четырех фундаментальных процессов.
Хотя диссипация свободной энергии, безусловно, является первым необходимым аспектом жизни, мы утверждаем, что оно должно сопровождаться тремя другими процессами — автокатализом, гомеостазом и обучением — и тогда описание жизненного процесса будет полным.
Это и есть 4 базовых принципа из заголовка статьи и их подробное описание выглядит так:
Диссипация — Y-жизнь не может существовать в равновесии. Второй закон термодинамики, при наличии механизмов передачи свободной энергии, позволяет соединять экзергонические реакции с эндергоническими, что необходимо для организации Y-жизни.
Используя массив наноразмерных молекулярных машин, жизнь рассеивает внешние химические неравновесия и/или преобразует низкоэнтропийные фотоны в высокоэнтропийное отработанное тепло, преобразуя одни неравновесия в другие (например, эндергонически зарождающиеся протонные градиенты и реакцию [АТФ]/[АДФ]). Для выполнения полезной работы жизнь преобразует АТФ→АДФ+H3PO4, который рассеивает неравновесие [АТФ]/[АДФ] [28 ,29 ].
Автокатализ — способность системы демонстрировать экспоненциальный рост репрезентативных показателей численности или популяции в идеальных условиях. Свойство автокатализа может проявляться в различных формах — включая самокатализ, кросс-катализ, сетевой автокатализ — до тех пор, пока эффект приводит к экспоненциальному росту подходящей метрики в идеальных условиях.
Культивированная система микроорганизмов демонстрирует автокаталитический рост популяции за счет репликации клеток в условиях изобилия ресурсов.
Гомеостаз — способность системы поддерживать ключевые внутренние переменные в рамках идеальных заданных значений. В динамическом мире возмущений и в сочетании с экспоненциальным ростом, описанным выше, Y-живая система должна иметь возможность ограничить изменения своих внутренних систем при изменении внешних условий.
Жизнь поддерживает гомеостаз при помощи сетей сенсоров, рецепторов и эффекторов. Вещество, регулируемое гомеостазом (например, ионы кальция), обычно связывается с рецепторами и способствует высвобождению каскада других веществ (например, гормонов). Эти цепочки соединений затем стимулируют соответствующий механизм реакции, чтобы вернуть уровень вещества в желаемое окно.
Хотя эти четыре принципа Y-жизни являются производными от наблюдений за жизнью в том виде, в котором мы ее знаем, новое определение намного более расширяемое. Эти четыре принципа представляют собой необходимые и достаточные требования к Y-живому состоянию, оставаясь при этом отделёнными от конкретных компонентов, составляющих систему.
Это иллюстрируется тем фактом, что существует множество систем, которые выполняют те же самые принципы, но достаточно отличаются друг от друга по форме (обсуждается далее). Следовательно, универсальность термина Y-жизнь вытекает из разумного ожидания того, что он может быть применен к еще неизведанным (или не изобретенным) системам, существующим в бесчисленных масштабах по всей Вселенной.
Может даже существовать класс систем, еще не открытых и не описанных, которые соответствуют всем четырём принципам Y-жизни, и, в дополнение, соответствуют некоторому пятому. Такие системы могут считаться Y-сверхживыми. Хотя открытие Y-сверхжизни, безусловно, стало бы началом новой научной парадигмы, мы пока что сохраняем агностицизм в отношении его существования.
Мы хотим подчеркнуть, что наше определение Y-жизни применяется на системном уровне. Принципы, которые могут быть применены к некой системе, зависят, в частности, от границ, которыми мы обозначаем эту систему. Например, какие принципы выполняют вирусы? Один вирус по отдельности не соответствует ни одному из принципов.
Вирусы в системе, состоящей из вирусов, бактерий и питательных веществ, могут совершать автокатализ и, путём разрушения своих жертв, диссипировать.
Вирусы в биосфере в целом, могут совершать не только автокатализ и диссипацию, но и обучаться (в процессе эволюции).
В некоторых экосистемах вирусы могут даже придавать системе гомеостатические атрибуты, вводя клеткам-хостам вспомогательные метаболические гены и поглощая органическое вещество через лизис [45 ].
Таким образом, аргумент о том, жив ли вирус, становится неактуальным при предположении, что «живость» возникает не на молекулярном, клеточном или организменном уровне. Как и некоторые другие исследователи [46 ,47 ], мы утверждаем, что живое состояние лучше всего оценивать в экосистемном или планетарном масштабе.
В качестве примера того, как один из принципов проявляется на системном уровне, рассмотрим обучение. Можно предположить, что скорость обучения естественным образом применима к отдельным видам, но, на наш взгляд, она более эффективно интерпретируется на системном уровне. Это происходит потому, что каждый вид развивается в согласии с другими видами и их абиотической средой. Следовательно, когда один вид учится, другие виды в системе, которые не вымерли, также должны учиться. Несмотря на то, что люди научились очень многому, другие виды, с которыми мы соседствуем на планете, учатся справляться с последствиями и изменениями, вызванными нашим обучением (например, грибковые виды учатся разрушать пластмассы [48 ,49 ,50 ] или люди учатся противодействовать патогенным микроорганизмам с помощью антибиотиков, которые затем в ответ обучаются резистентности [51 ]).
Мы также отмечаем, что важность и актуальность этих принципов отделены от той легкости, с которой мы можем измерить их присутствие.
Для примера, измерение автокаталитического свойства, скорее всего, будет нетривиальным, если только рассматриваемые организмы/системы не могут быть культивированы. В частности, получения биосигнатур с удаленных экзопланет, вероятно, будет недостаточно для выявления автокаталитических свойств, если только эти сигнатуры не собираются в течение очень длительных периодов времени.
Следовательно, для этого принципа существует разделение между значимостью (это свойство имеет основополагающее значение для определения Y-жизни) и легкостью измерения (его может быть нелегко оценить).
Однако иногда автокаталитическое свойство проявляется очень драматично. Пока мы пишем эту статью, мир переворачивается с ног на голову сущностью, которая одновременно считается неживой и состоит только из небольшого генома РНК и набора белков. Каждая диаграмма, которая отслеживала COVID-19, показывала экспоненциальный рост (автокатализ) в первой фазе, демонстрируя как крошечные биологические сущности могут показать экстремальные нелинейные динамические изменения в короткое время.
Y-субжизнь
Помимо предоставления простого «контрольного перечня» критериев для определения того, является ли динамическая система живой или нет, эти четыре принципа также позволяют нам рассмотреть Y-жизнь в контексте других явлений во Вселенной.
Мы определяем Y-жизнь как любую систему, которая выполняет все четыре столба, и Y-субжизнь как любую систему, которая выполняет некоторые, но не все эти функции (рис. 5).
Только диссипация — тепловая диффузия или любой термодинамически необратимый процесс.
Только гомеостаз — идеальный газ в равновесии. Изолированная система, подобная этой, всегда возвращается в состояние равновесия после внутренних или внешних колебаний.
Диссипация и автокатализ — огонь — часто обсуждаемый пример диссипации и автокатализа. Он демонстрирует гомеостаз определенных переменных (например, температура горения естественным образом не выходит за некоторые пределы), но его неспособность полностью регулировать свое состояние или учиться на опыте удерживает его в неживом мире. Другим значимым примером может служить экспоненциальный рост продуктов в нелинейных химических реакциях (например, в реакции Бутлерова).
Диссипация и гомеостаз — затухающий гармонический осциллятор преобразует кинетическую энергию в тепловую и всегда возвращается в положение равновесия.
Диссипация и обучение — искусственная нейронная сеть — это пример системы, которая диссипативна и может обучаться, но не обязательно демонстрирует автокаталитический рост или гомеостаз (например, она сама по себе не поддерживает температуру собственного аппаратного обеспечения). Можно утверждать, что полезность нейросетей заставляет нас плодить их с экспоненциальной скоростью, но это совсем другая дискуссия.
Рассеивание, автокатализ и обучение — живая система, которая попала в ловушку трагедии общин. Примерами могут служить привнесенные на остров инвазивные виды, которые уничтожают источники пищи настолько быстро, что источники пищи более не подлежат восстановлению.
В качестве другого примера можно также привести антропогенное изменение климата. Обратите внимание, что эти случаи в решающей степени зависят от того, где проводится граница системы (например, при включении или не включении человека).
В действительности, такая форма субжизни или Y-субжизни менее вероятна, потому что если система способна учиться, то в принципе она может научиться регулировать себя гомеостатически (если может учиться достаточно быстро).
Рассеивание, гомеостаз и обучение — «умный» домашний термостат, который следит за поведением жильцов с течением времени. Эта система не может воспроизводить себя, но потребляет свободную энергию, способна к примитивному обучению и может регулировать собственную локальную температуру.
Рассеивание, автокатализ и гомеостаз — например, реакция Белоусова-Жаботинского. Показано, что некоторые неравновесные химические реакции растут экспоненциально, а также способны регулировать собственную локальную температуру [52 ,53 ,54 ,55 ].
Все 4 — Y-жизнь (и жизнь, как её подмножество).
Что касается гомеостаза в системах равновесия (область 2 на Рис. 5), то здесь есть некоторые тонкости. Наше утверждение о том, что гомеостаз происходит в изолированных системах, заключается просто в том, что они являются архетипами устойчивости (по определению). Однако этот вопрос может быть спорным. Можно утверждать, что в момент флуктуации происходит мгновенное создание свободной энергии. Однако, использование такого колебания потребовало бы измерений и обработки информации, и, как показано в [56 ], для этого в системе с ограниченным количеством памяти потребуется стирание, которое не может быть сделано бесплатно, а значит, свободной энергии, создаваемой колебаниями системы равновесия, на самом деле не существует.
В целом любое возмущение такой системы исчезнет за конечное время по мере уравновешивания системы, поэтому мы допускаем гомеостаз в системах равновесия (однако, биологические гомеостатические процессы, как правило, происходят за счет потребления свободной энергии).
Существует несколько примечательных макромолекулярных комплексов, которые являются как диссипативными, так и автокаталитическими, но не обязательно гомеостатическими (области 3 и 6 на рис. 5).
Выше мы уже обсуждали вирусы, которые являются белково-геномными комплексами. Прионы являются, по сути, пептидными конформационными вирусами, так как они размножаются через патологическое распространение собственной конформации на существующие пептиды (различных конформаций) [57 ,58 ,59 ].
Существуют также автокаталитические гены, известные как транспозоны: «транспонируемые или подвижные элементы, способные к паразитоподобной пролиферации в геноме хозяина» [60 ,61 ,62 ,63 ].
Например, интернет-мемы явно являются автокаталитическими и диссипативными (учитывая затраты на вычислительную энергию, связанные с их распространением и коммуникацией). Кроме того, система «участие в социальных медиа плюс мемы» учится коллективно, и тем самым оказывается в регионе 6 на рис. 5.
Кроме того, по аналогии с биологическими вирусами, фейковые новости, которые лишь слабо коррелируют с реальностью, могут усиливаться различными эффектами до такой степени, что вызывают разрушительные социально-политические последствия. (Прим. переводчика — это одна из двух основных причин, почему я решил перевести данную статью).
В этой главе мы представили список все более Y-жизнеподобных явлений, которые могут создавать иллюзию того, что происхождение Y-жизни всегда происходит простым, поэтапным образом — т.е. пребиотическая диссипативная структура должна сначала демонстрировать экспоненциальный рост (возможно, репликацию), приобретать гомеостатические регуляторные механизмы, а затем, наконец, учиться.
На наш взгляд, вероятно также, что относительно простые системы, способные к рудиментарной обработке информации, могут возникнуть изначально и что способность этих систем со временем оптимизировать свои диссипативные, автокаталитические и гомеостатические черты определит их конечную судьбу (см. также [66 ]).
Например, первая жизнь на Земле почти наверняка не использовала ДНК для хранения информации или какие-либо узнаваемые ферменты в своей метаболической сети. После бесчисленных колебаний между случайностью и необходимостью эволюция всё же породила знакомые нам макромолекулы, которые мы наблюдаем сегодня. Далее мы покажем, как понятие «Y-жизнь» может изменить наши подходы к исследованию истоков жизни.
Y-жизнь и исследования происхождения жизни
Что касается сценариев происхождения жизни, Y-жизнь включает в себя любую систему, которая удовлетворяет четырем принципам, описанным выше, но может выполнять три классические характерные функции — репликацию, метаболизм и компартментализацию — используя компоненты, которые недоступны для земной жизни.
Идея о том, что эти три характерные функции определяют необходимые и достаточные условия жизни, была подробно теоретически исследована в области создания искусственной жизни и концепций аутопоэзис [67 ] и «Хемотон» [68 ].
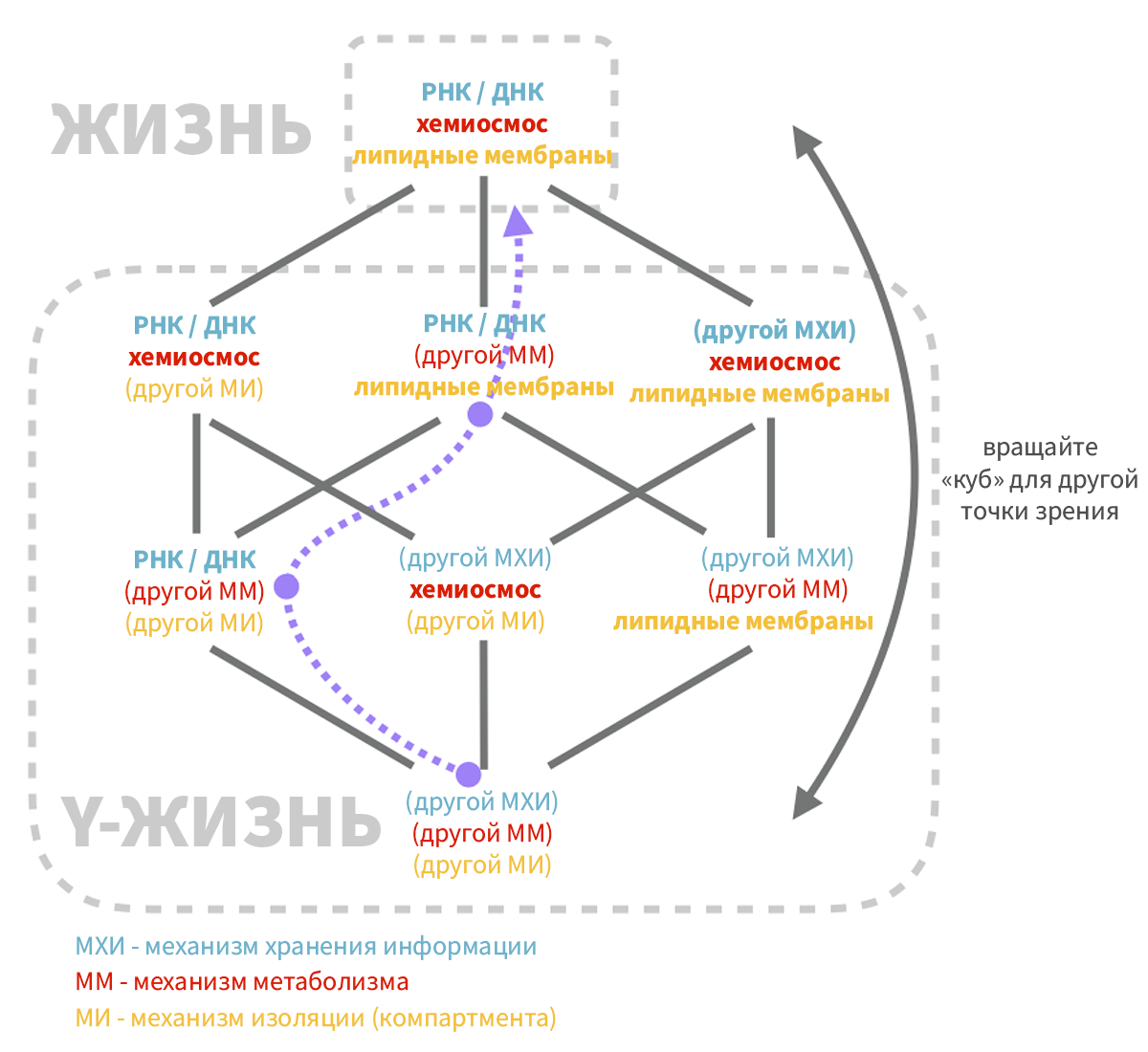 Рисунок 6.
Рисунок 6.
На рисунке 6 показан «куб», вершины которого представляют живые системы с различными комбинациями компонентов, выполняющих три характерные функции. На одной вершине жизнь выполняет эти характерные функции, используя механизмы РНК/ДНК, хемиосмоса и липидных мембран. Вы можете удалиться от земной жизни на 1, 2 или 3 шага по ребрам этого куба и получить самые различные комбинации.
Как обсуждалось ранее, довольно трудно распознать компоненты, которые использовались в самых ранних формах жизни имея на руках только информацию о сильно развитых и сложных механизмах, которые используются жизнь сегодня. Мы можем предложить только две метафорические аналогии, которые полезны в этом вопросе. Первая — это поразительные различия в составе и структуре между строительными лесами и готовым зданием. Вторая — это аналогия между биологической эволюцией и историей локомотивов (рис. 7).
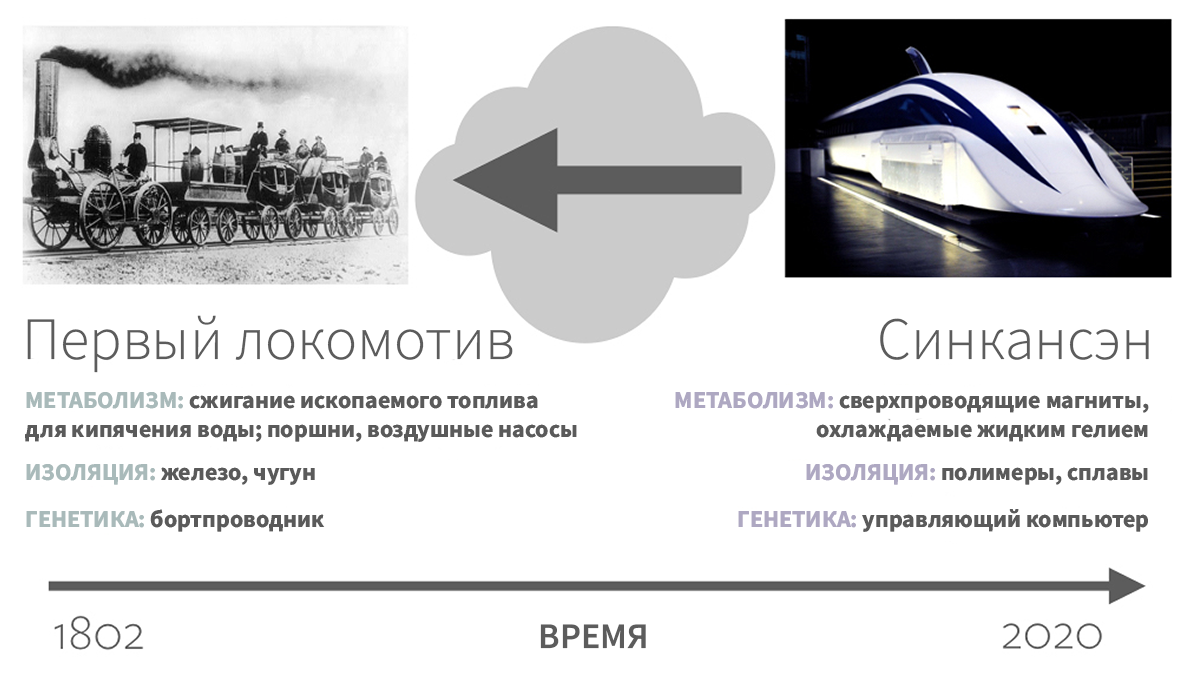 Рисунок 7.
Рисунок 7.
Рассмотрим сверхсовременный японский поезд Синкансэн (с «метаболизмом», основанном на сверхпроводящих магнитах, охлаждаемых жидким гелием, «компартментализацией» из композиционных материалов, таких как углеродное волокно, полимеры и сплавы, и «обработкой информации» с помощью системы компьютерного управления). Изучая Синкансэн и ничего больше не зная, можно ли определить, как выглядел первый локомотив («метаболизм», осуществляемый при сжигании ископаемого топлива, «компартментализация» из чугуна и древесины, и «обработка информации» в виде бортпроводника)?
Учитывая, что компоненты локомотива могут взаимозаменяться между собой человеческими руками, эта аналогия может показаться неуместной с точки зрения общепринятого представления о биологической эволюции как о постепенном процессе, движущей силой которого является естественный отбор микроулучшений к уже существующим элементам. Например, в так называемых «луковых гипотезах» об эволюции жизни [69 ,70 ,71 ] предполагается, что жизнь начиналась с метаболического ядра реакций, вокруг которого складывались все дальнейшие слои сложности.
Вполне возможно, что на самых ранних этапах жизни такие переходы были лавинными: примитивные живые системы боролись за господство в относительно плоском ландшафте приспособляемости, легко обменивались компонентами посредством горизонтальных передач информации и материала. С изобретением механизма, который давал большее селективное преимущество, чем остальные, этот механизм — будь то рибосома или АТФ-синтез — стал бы стабильным аттрактором в эволюционной траектории земной биологии.
По нашему мнению, истина, вероятно, наиболее близка к гибриду этих «веревочных» и «луковых» концепций. Появление жизни и ранние стадии, вероятно, хорошо представлены концепцией веревки, в которой различные волокна соединяются, вносят свой вклад и покидают веревку. Основываясь на аналогии с «длинной веревкой», мы предполагаем, что «поглощения» Кэрн-Смита могли быть скорее внезапными, чем постепенными.
Однако, как только эволюция наткнулась на волокно с превосходной эффективностью или стабильностью, эта особенность быстро интегрируется и становится центральной нитью веревки жизни, навсегда укореняясь в молекулярном ядре биосферы. Слои функций, накапливающиеся вокруг центральной нити веревки, предотвращали бы потерю основных компонентов из веревки. Это приводит к «луковичной» истории, которую можно разобрать на части с помощью современных методов, таких как молекулярная филогения и метаболомика.
Обратите внимание, что «фиксация» функциональности происходит во многих сложных системах, включая экономические, технологические и политические. Системные ядра, которые взаимодействуют со многими более высокоуровневыми подсистемами, становятся настолько функционально связанными с различными модулями, что любые изменения в ядре будут вредными или катастрофическими для согласованного целого.
Это может быть верно, даже если существуют более совершенные системы-кандидаты на создание ядра. Например, многие европейские города в течение долгого времени росли органически и теперь имеют несколько фрактальную организацию. Сравните это с гораздо более спланированными сетчатыми структурами американских городов. Изменение фундаментальной структуры европейских городов в настоящее время слишком дорогостоящее, поэтому они надолго застряли с их спонтанными планировками.
Также вполне вероятно, что современные компоненты основных биохимических систем изначально выполняли различные функции. Биосфера изобилует примерами такой экзаптации: смена функции или кооптация компонента для другого использования. Перья изначально давали тепло и сигнальные способности и только позже стали орудиями полета. Крылья, вероятно, использовались для увеличения скорости бега прежде чем стали использоваться для полета [74 ]. Водный окислительный комплекс, возникший в эпоху кислородного фотосинтеза, возможно, изначально использовался для окисления марганца [75 ]. Дальнейшие примеры экзаптации смотрите в работе [76 ].
Таким образом, мы должны быть осторожны в попытках объявления однозначной универсальности того или иного компонента в современной биосфере и его необходимости для появлении жизни.
Возможно, длинные нити нуклеотидов не были первой системой обработки информации. Возможно, полифосфатные цепи не были первой «энергетической валютой» жизни. Возможно, первые мембраны не состояли из органических углеводородов.
Сценарии происхождения жизни, которые ищут абиотические пути синтеза специфических «строительных блоков» жизни, основываются на предположении, что эти молекулы: (1) стояли у истоков жизни; (2) выполняли те же функции в истоках жизни, что и сегодня; (3) проявляли функциональность сразу после формирования, т.е. после синтеза и начала применения всех компонентов жизни их способность к сложной обработке информации просто взяла и появилась (некоторые даже описывали это третье предположение как форму современного «витализма» [77 ]). Этот узкий подход неизбежно слеп к сценариям, в которых жизнь могла бы начинаться с использования альтернативных компонентов, как это могло бы произойти не только на Земле, но и в других местах Вселенной.
Итак, исследуя возникновение жизни, что же мы ищем, если не конкретные биомолекулы? Определение Y-жизни дает ответ: мы ищем систему, которая демонстрирует диссипацию, автокатализ, гомеостаз и обучение. Основываясь на общих процессах, а не на конкретных компонентах, выполняющих конкретные задачи, мы открываем свой разум для исследования всех систем, которые отображают эти возникающие свойства, освобождая себя от ограничений точных химических рецептов, рецепты которых содержат предположения, которые могут ограничить наши исследования появления во вселенной поведения, похожего на жизнь.
Какой может быть Y-жизнь? Примеры альтернативных компонентов в гипотезах происхождения жизни
Сообщество исследователей происхождения жизни выдвигает множество гипотез, которые предполагают поэтапное возникновение через Y-жизнеподобные фазы. Используя нашу классификацию, эти возникающие системы можно было бы классифицировать как Y-живые или Y-субживые, потому что они используют радикально отличные от современной жизни компоненты для достижения одного или более из базовых принципов жизни. Данная глава описывает несколько важных примеров гипотез происхождения, которые используют альтернативные компоненты.
В гипотезах возникновения, затрагивающих гидротермальные системы, компартментализация достигается не за счет самопроизвольной компоновки липидных мембран, а за счет лабиринта неорганических поровых пространств минералов. Только после изобретения биосинтеза липидов и последующего изобретения биохимии клеточных стенок появились живые клетки, выходящие из гидротермальных границ [95 ]. Однако до того, как они стали свободноживущими прокариотами, сложные химические сети, размещённые внутри их минеральных оболочек, все еще можно было считать Y-живыми (или Y-субживыми до достижения всех четырех базовых принципов).
Y-Жизнь на Титане
Биохимия жизни — от её водной природы до сильной зависимости от атомов C H N O P S — является отражением физических и химических условий Земли. Другие миры, в которых имеются потенциально схожие места обитания, например, гидротермальные системы в Европе или Энцеладе, могут быть населены живыми существами, биохимически сходными с жизнью. Однако в мирах, которые занимают радикально иные физические и химические пространства, чем на Земле, любая существующая экзобиология, несомненно, будет Y-жизнью, а не жизнью.
Одной из интригующих возможностей является Титан, единственный другой мир в Солнечной системе, который, как известно, обладает на своей поверхности устойчивыми озёрами жидкости. Несмотря на широкое сходство Титана с Землей (атмосферное давление, геоморфология, активный метановый цикл, схожий с гидрологическим циклом Земли и т.д.), если бы экзобиология существовала на Титане, она возникла бы и развивалась в среде, невероятно непохожей на Землю. К основным отличиям окружающей среды относятся: крайне низкие поверхностные температуры ∼94 K (что приводит к экзотической органической химии, основанной на силах Ван-дер-Ваальса [100 ,101 ]); недостаток кислорода для биохимии (что приводит к гипотезам для N-замещенных биохимий [102 ]); и неполярный растворитель CH4-C2H6 (что приводит к догадкам об альтернативных мембранных структурах).
Механотрофы
В духе дальнейших размышлений о жизни, какой мы ее не знаем, мы представляем вам гипотетическую форму жизни, которая использует альтернативную диссипативную/метаболическую систему — преобразование механической работы в химическое неравновесие. Земная жизнь использует различные внешние источники свободной энергии, от окислительно-восстановительных пар до солнечного излучения и градиентов плотности протонов и даже электронов (например, электротрофы [106 ]). Известно, что макроскопические организмы эксплуатируют абиотические источники механической работы и используют их в своих интересах; в качестве примера можно привести наезд лосося на турбулентные вихри для плавания вверх по течению [107 ,108 ], птиц, улавливающих восходящие потоки воздуха для взлёта на большие высоты, а также строительство человеком ветряных турбин и гидроэлектростанций. Однако каталогизированного примера организма, преобразующего механическую работу непосредственно в свой метаболизм, не существует.
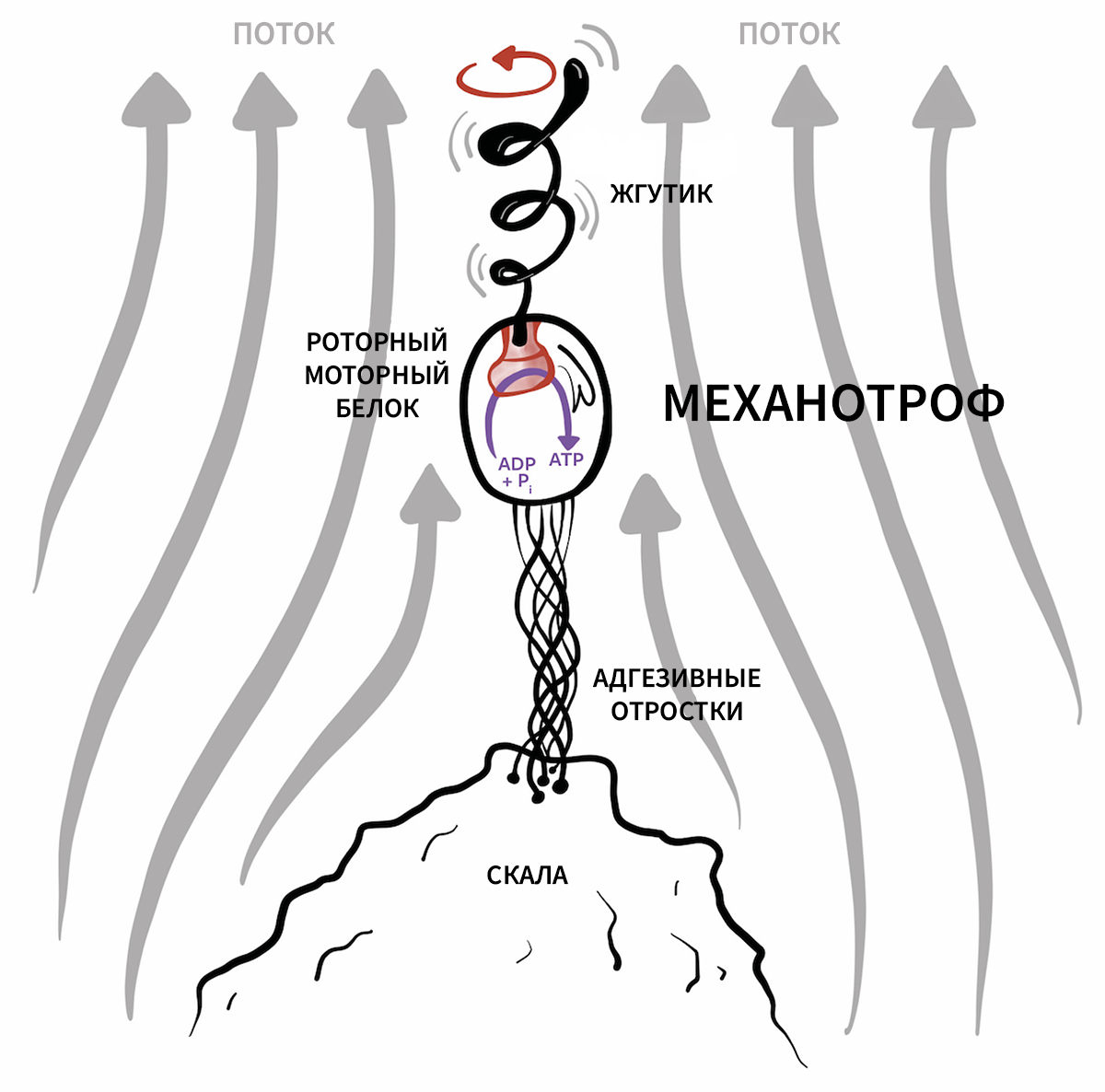 Рисунок 8.
Рисунок 8.
Почему этот процесс не может быть обращен вспять для питания механотрофного организма? Представьте себе одноклеточный организм в проточной воде, прикрепленный к скале через тросоподобную нить. Предположим, что такой организм оснащен типом жгутиковых моторных белков, которые были изучены у плавающих бактерий или одноклеточных эукариот. Поток воды вызывает вращательное движение жгутиковых нитей, и это вращательное движение используется для синтеза АТФ (или, возможно, какого-либо другого эндергонического химического шага в более экзотической Y-жизни), как показано на рис. 8. Реально ли это, зависит от термодинамических и гидродинамических условий.
Обратите внимание, что такой механотрофный организм может иметь конструкцию, отличающуюся от той, которую архетипичная бактерия использует для плавания. Например, он может использовать жёсткий к скручиванию трос, прикрепленный к молекуле молекулярного генератора (жгутик приводится в движение), и вся клетка может вращаться вокруг комплекса трос-генератор. В этом случае жгутик фиксируется относительно тела ячейки (на противоположном конце тросо-генераторного комплекса) и вместе с ячейкой вращается.
Гидродинамические детали, эффективность и энергия такого организма будут детально изучены в будущей работе. Здесь мы просто хотим подчеркнуть возможность существования такого организма. Это был бы наглядный пример Y-жизненной формы, хотя не исключено, что такой организм существует на Земле, но до сих пор просто остался незамеченным.
Заключение (от переводчика)
Все источники вы можете увидеть в оригинальной статье. Я не стал полностью убирать ссылки на них в переводе, так как это затрудняло корректную передачу смысла. Авторы очень многословны, поэтому некоторые, незначительные для общего сюжета абзацы я не стал включать в текст.
Я перевёл эту статью, так как она может помочь взглянуть по новому на некоторые социально-экономические явления, а так же помочь исследователям в области машинного обучения (коим я сам являюсь) нащупать путь к созданию сильного искусственного интеллекта.
Источник